熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
摘要
3-亞磺酰丙酰輔酶A(3SP-CoA)脫亞磺酰酶(Acd_DPN7;EC 3.13.1.4)是在研究β-變形菌門Advenella mimigardefordensis菌株DPN7T中3,3'-二硫代二丙酸(DTDP)分解代謝途徑時發現的。DTDP是一種有機二硫化物,是細菌合成聚硫酯(PTE)的前體,在生物技術PTE生產中具有重要意義。Acd_DPN7催化從3SP-CoA中提取硫,這是DTDP分解代謝過程中的一個關鍵步驟。本文展示了分辨率為1.89?的脫輔基Acd_DPN7晶體結構,以及分辨率為2.30?的其與底物類似物琥珀酰輔酶A的輔酶A部分形成的復合物晶體結構。脫輔基結構顯示,Acd_DPN7屬于酰基輔酶A脫氫酶超家族折疊,它是一個四聚體,每個亞基含有一個黃素腺嘌呤二核苷酸(FAD)分子。該酶不顯示任何脫氫酶活性。脫氫酶活性需要在246位或366位有一個催化堿基(Glu或Asp殘基),而在這個脫亞磺酰酶中,這兩個位置分別是一個谷氨酰胺和一個甘氨酸。輔酶A在晶體復合物中的定位使得包含3SP-CoA的底物復合物建模成為可能。這表明Arg84是脫亞磺酰反應中的一個關鍵殘基。Arg84Lys突變體顯示出完全喪失酶活性,表明精氨酸的胍基對脫亞磺酰作用至關重要。Acd_DPN7是首個報道的具有酰基輔酶A脫氫酶折疊的脫亞磺酰酶,這突顯了這種酶支架的多功能性。
1.引言
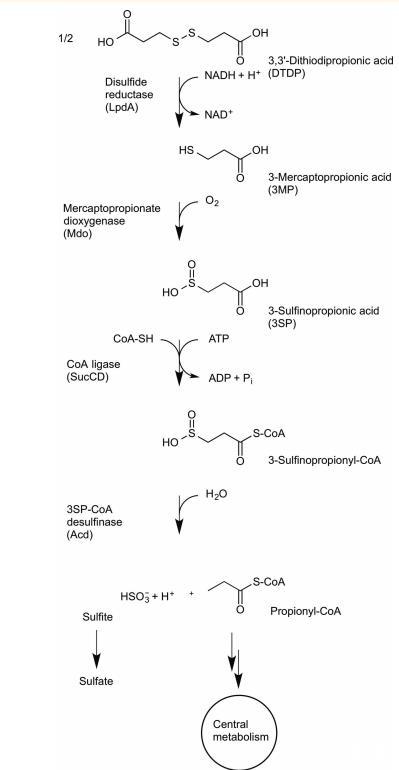
圖1 DTDP 的降解過程。二硫鍵化合物3,3‘-二硫代二丙酸(DTDP)通過NADH依賴性二硫鍵還原酶(Wu¨bbeler等,2010)裂解為兩個3-巰基丙酸(3MP)分子。3MP經雙加氧酶(Mdo;Bruland等,2009)氧化生成3-亞磺酰丙酸(3SP)后,琥珀酸-CoA連接酶(SucCD;Schu¨rmann等,2011)催化其活化為3SP-CoA。隨后,3SP-CoA被亞磺酸脫氫酶(Acd)催化水解裂解,生成丙酰輔酶A和亞硫酸鹽。丙酰輔酶A最終通過甲基檸檬酸循環進入中心代謝途徑,而亞硫酸鹽很可能被亞硫酸鹽氧化酶氧化為硫酸鹽。
3-亞磺酰丙酰輔酶A(3SP-CoA)降解為亞硫酸鹽和丙酰輔酶A是3,3'-二硫代二丙酸(DTDP)細菌分解代謝途徑的最后一步(圖1)。DTDP是一種有機二硫化物,是聚硫酯(PTE)合成的前體。為了提供一種工程化適合生物技術PTE生產菌株的策略,對Advenella mimigardefordensis菌株DPN7T中DTDP降解途徑的闡明以及所涉及基因的鑒定已經完成。最近,有報道稱在β-變形菌門Advenella mimigardefordensis菌株DPN7T的DTDP分解代謝過程中,分離出一種脫亞磺酰酶(EC 3.13.1.4),該酶催化從3SP-CoA中提取硫。純化的DTDP的降解。二硫化物3,3'-二硫代二丙酸(DTDP)被NADH依賴性二硫化物還原酶裂解成兩分子3-巰基丙酸(3MP)。在雙加氧酶(Mdo)將3MP氧化為3-亞磺酰丙酸(3SP)后,琥珀酰輔酶A連接酶(SucCD)催化其活化生成3SP-CoA。隨后,3SP-CoA脫亞磺酰酶(Acd)催化水解裂解,生成丙酰輔酶A和亞硫酸鹽。丙酰輔酶A最終通過甲基檸檬酸循環進入中心代謝,而亞硫酸鹽很可能被亞硫酸鹽氧化酶氧化為硫酸鹽。
蛋白質顯示出約150-200 kDa的天然分子量,表明其為同源四聚體結構,并且每個亞基含有一個非共價結合的黃素腺嘌呤二核苷酸(FAD)輔因子,這與酰基輔酶A脫氫酶超家族的其他成員一致。在之前的一項研究中,體外測定證實純化的酶將3SP-CoA轉化為丙酰輔酶A和亞硫酸鹽,但它無法進行脫氫反應,這是酰基輔酶A脫氫酶(Acd)超家族成員通常催化的反應。
最近,來自Variovorax paradoxus的Acd_TBEA6、來自Burkholderia xenovorans的Acd_LB400和來自Cupriavidus necator的Acd_N-1被鑒定為酰基輔酶A家族中的3SP-CoA脫亞磺酰酶,與Acd_DPN7的序列同一性約為60%。這三種Acd均未對任何測試的酰基輔酶A硫酯(即丁酰輔酶A、異丁酰輔酶A、戊酰輔酶A、異戊酰輔酶A、戊二酰輔酶A、琥珀酰輔酶A和3SP-CoA)進行脫氫。
除了這些新的脫亞磺酰酶外,迄今為止已報道了另外三種細菌酶,它們催化各種底物上的脫亞磺酰反應,產生亞硫酸鹽作為最終產物之一。一種是大腸桿菌的吡哆醛磷酸(PLP)依賴性半胱亞磺酸脫亞磺酰酶(CSD;EC 4.4.1.16),它具有硒代半胱氨酸裂解酶和半胱氨酸脫硫酶側活性。第二種是PLP依賴性天冬氨酸β-脫羧酶(EC 4.1.1.12),它可以接受半胱亞磺酸并催化脫亞磺酰作為副反應。第三種酶DszB(EC 3.13.1.3)催化從2'-羥基聯苯-2-亞磺酸鹽以及2-苯基苯亞磺酸鹽中水解亞磺酰基。已報道的脫亞磺酰酶均未顯示出與Acds(EC 1.3.8.x)的結構相似性。
Acds屬于黃素酶超家族,主要催化酰基輔酶A硫酯的α,β-脫氫,使用電子轉移黃素蛋白(ETF)作為其生理電子受體。Acds主要參與脂肪酸氧化或支鏈氨基酸代謝,但也參與不太常見的細菌代謝途徑。例如,來自Thauera aromatica的芐基琥珀酰輔酶A脫氫酶參與厭氧甲苯分解代謝途徑,以及參與乙基丙二酰輔酶A途徑的2-甲基琥珀酰輔酶A脫氫酶(Mcd)。
在此,我們報道了3SP-CoA脫亞磺酰酶Acd_DPN7的兩種晶體結構。分辨率為1.89?的脫輔基酶晶體結構揭示了典型的酰基輔酶A脫氫酶超家族折疊。更具體地說,Acd_DPN7是一個四聚體,每個單體含有一個非共價結合到多肽上的FAD。與琥珀酰輔酶A共結晶產生了含有輔酶A的全酶復合物。該結構的分辨率為2.30?,顯示了輔酶A的結合方式,并為底物結合提供了線索。晶體結構揭示了活性位點周圍殘基的構型,并表明Arg84在脫亞磺酰作用中起著重要作用。誘變顯示Arg84是必需的,結構比較提供了關于脫亞磺酰酶如何在脫氫酶超家族中出現的見解。
2.材料與方法
2.1.化學品
純度至少為95%(根據氣相色譜法)的3SP是使用先前描述的方法合成的,但通過重復雙-(2-羧乙基)砜中間體的堿裂解步驟進行了修改。3SP的合成和純度通過GC和GC/MS進行分析。用于酶測定的3SP-CoA(同樣不可商購)的原位形成是根據其他地方描述的方法進行的。其他輔酶A硫酯,即丙酰輔酶A、丁酰輔酶A、異丁酰輔酶A、戊酰輔酶A、異戊酰輔酶A、琥珀酰輔酶A和戊二酰輔酶A,是通過基于酸酐法的改良技術合成的。這種改良方法先前已有描述。
 相關新聞
相關新聞