熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
由于脫氧肌紅蛋白向氧合肌紅蛋白的轉化是鐵依賴性的,并可能受細胞氧化還原狀態的影響,使用CM-H2DCFDA測量了經過氧氣轉運測定預處理后選定菌株的氧化還原狀態。CM-H2DCFDA產生的熒光強度在氧氣轉運測定預處理后的酵母菌株之間沒有顯著差異。這表明在氧氣轉運測定之前,模擬、HsAQP1、NtPIP1;3和AtPIP1;2菌株中的細胞氧化還原狀態相似。與早期報告類似,在NtPIP1;2菌株中檢測到過氧化氫通透性增加。然而,在NtPIP1;3、HsAQP1或模擬菌株中未測量到過氧化氫通透性增加,而在AtPIP1;2菌株中觀察到略高的熒光強度(提示過氧化氫通透性增加)在統計學上不顯著。
酵母氧氣消耗能力。異源表達NtPIP1;3和HsAQP1的酵母細胞分別顯示出比模擬對照高2.3倍和1.8倍的氧氣吸收速率,并且從溶液中消耗氧氣的速度顯著更快。表達AtPIP1;2的酵母細胞的氧氣吸收速率以及從溶液中耗盡氧氣的時間與模擬對照相比沒有顯著差異。異源表達水通道蛋白對酵母細胞直徑沒有顯著影響,在模擬、HsAQP1、NtPIP1;3和AtPIP1;2菌株中測量值分別為2.87±0.05、2.74±0.06、2.98±0.06和2.87±0.08微米(平均值,n=50±標準誤)。
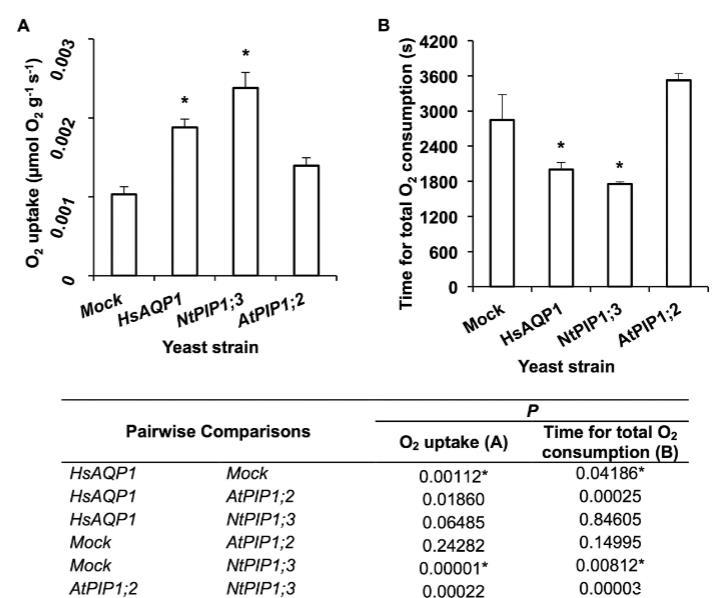
圖5.酵母菌株在1000秒內的氧氣消耗情況。(A)HsAQP1、NtPIP1;3、AtPIP1;2和模擬菌株(對照)的呼吸速率。(B)HsAQP1、NtPIP1;3、AtPIP1;2和模擬菌株(對照)的氧氣完全消耗時間。在向經氮氣鼓泡的SD-L-U+葡萄糖培養基中的酵母懸浮液供應空氣,使其可溶性氧氣濃度達到飽和濃度235μmol L-1后,立即使用氧氣微傳感器監測并每秒記錄酵母懸浮液中氧氣濃度的下降。星號表示與模擬菌株存在統計學顯著差異(下表顯示P值)(ANOVA,Tukey檢驗,P≤0.05,n=6±標準誤)。
低氧條件下煙草根中PIP轉錄豐度和ATP水平。我們檢測了在礦物溶液培養中遭受淹水誘導低氧處理兩天和七天的煙草植物的轉錄水平。經過兩天低氧處理后,顯示高氧氣轉運速率的水通道蛋白NtPIP1;3在葉片和根中的轉錄水平比通氣良好的植物增加了約四倍。在通氣良好的植物中,葉片和根中的水通道蛋白轉錄水平在第二天和第七天保持相似。在葉片中NtPIP1;4以及根中NtPIP1;1和NtPIP1;2的轉錄水平也測量到相對較小的增加。七天后,在低氧葉片和根中測量到NtPIP1;3的急劇增加(分別比通氣對照高約12倍和22倍)。葉片中NtPIP2;1也有約三倍的增加。在低氧處理下,從第二天到第七天,葉片和根中的NtPIP1;3轉錄水平急劇增加,同時根尖段的ATP水平顯著增加。處理七天后,每個根段中低氧和通氣良好的根具有相似的ATP水平。低氧植物外觀健康翠綠,沒有出現缺氧癥或其他可見的氧氣缺乏跡象。
討論
在本研究中,我們通過測量肌紅蛋白峰值波長附近吸光度隨時間的變化,研究了水通道蛋白對酵母完整細胞和酵母原生質體跨膜氧氣轉運的潛在貢獻。在完整細胞測定中,A541在前60秒內增加,這使我們能夠篩選表達推定的氧氣轉運水通道蛋白的菌株。A541的變化可能代表幾個過程的組合,包括氧氣擴散、脫氧肌紅蛋白的氧合、氧合肌紅蛋白和高鐵肌紅蛋白之間的轉化以及氧氣消耗。細胞壁的存在可能會阻礙肌紅蛋白吸光度的變化,并在初步篩選中導致對氧氣擴散的低估。此外,在混合酵母懸浮液和通氣緩沖液期間可能會對吸光度讀數造成假象。酵母原生質體測定旨在通過眾多預防措施和更多重復來消除這些潛在缺陷,并最大化肌紅蛋白氧合的信號。結果表明,酵母原生質體測定中的ΔA319/ΔA341也可能是測量氧氣通過水通道蛋白轉運的高度敏感參數。
我們發現能通過促進氧氣通道來增強肌紅蛋白氧合的人水通道蛋白HsAQP1,也曾被報道當在非洲爪蟾卵母細胞中異源表達時能促進二氧化碳轉運。然而,其他主要的二氧化碳轉運水通道蛋白,包括AtPIP1;2、NtPIP1;2和LbAQP1,在酵母中表達時并未促進氧氣轉運。這表明水通道蛋白直系同源物已經發展出一定程度的氧氣轉運特異性。對所有分析的20種水通道蛋白的比對并未顯示氧氣轉運水通道蛋白獨有的共有殘基。看來保守殘基是物種依賴性的,而非與轉運能力相關。然而,值得注意的是,所有六種氧氣轉運水通道蛋白在物種間具有高度保守的29個氨基酸殘基,包括大多數Asn-Pro-Ala簽名基序和Ar/R選擇性過濾器的殘基。在NtPIP1;3中,通常位于第二個NPA基序中的天冬酰胺殘基被蘇氨酸(Thr-235)取代,這可能與其高度增強的氧氣轉運能力潛在相關。
對疏水氣體(氧氣、二氧化碳和一氧化氮)滲透的計算一致顯示通過水孔的能量屏障值相似,為5-6千卡每摩爾。人類HsAQP1四聚體的膜蛋白模擬系統證明了在四個單體之間中心存在一個孔,該孔主要由疏水殘基排列,可能參與氣體而非水的轉運。可以推測,該孔的存在及其精確結構賦予了不同水通道蛋白氣體轉運特異性。如果證明正確,該孔的門控特性可以作為靶點來改變跨膜氣體通過的速率。
酵母氧氣吸收速率的結果證實了氧氣轉運測定的結果,指出了孔介導的轉運對呼吸作用的重要性。癌細胞中經常報道HsAQP1轉錄水平增加,有時也伴隨其他水通道蛋白的增加,HsAQP1的表達水平通常與細胞生長、腫瘤分級和轉移相關。也有報道稱,刪除HsAQP1能有效減小乳腺腫瘤大小和肺轉移,并且HsAQP1沉默抑制了骨肉瘤細胞的增殖和侵襲性。雖然對HsAQP1表達與癌性生長之間聯系提出的解釋主要集中在水轉運上,但也應考慮快速生長的癌細胞對氧氣的高需求與HsAQP1促進氧氣轉運之間的關聯。
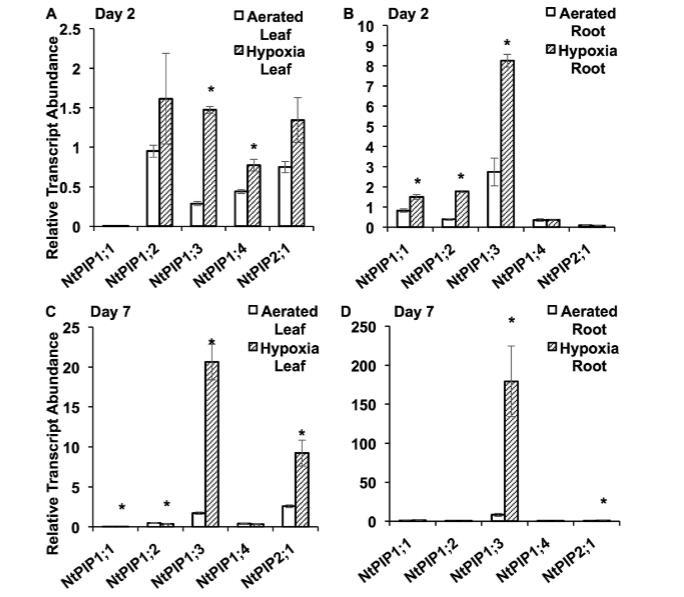
圖6.暴露于通氣良好和低氧條件后,煙草質膜內在蛋白的轉錄豐度。(A)暴露2天后葉片中和(B)根中,以及(C)暴露7天后葉片中和(D)根中所選PIPs的相對轉錄豐度。PIPs的轉錄豐度通過qRT-PCR分析中的標準曲線法測定,并針對兩個參考基因EF1-α和L25轉錄豐度的幾何平均值進行標準化。星號表示同一天通氣良好與低氧處理之間基因表達的顯著差異;下表列出了第2天與第7天之間比較的P值(ANOVA,Tukey檢驗,P≤0.05,n=6±標準誤)。
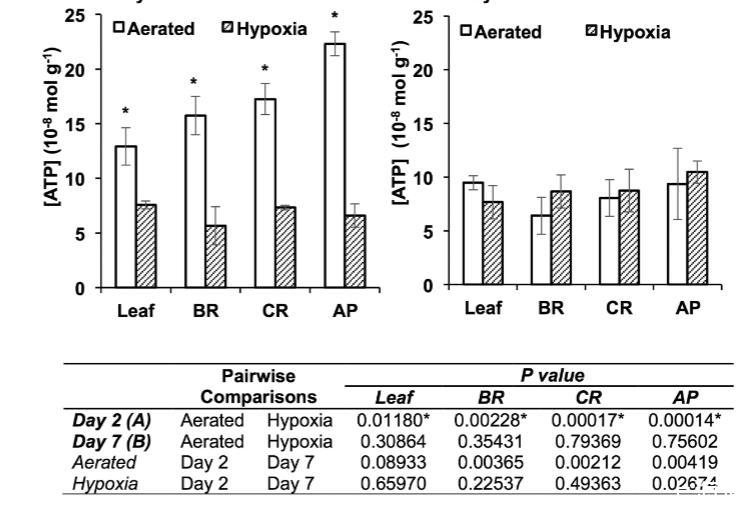
圖7.遭受低氧和通氣良好條件的煙草植株葉片以及根基部、根中部和根尖段的ATP水平。(A)處理2天后的ATP水平。(B)處理7天后的ATP水平。ATP水平通過檢測螢光素酶/螢光素反應中的生物發光信號來確定。星號表示同一天同一組織中通氣良好與低氧處理之間ATP水平的顯著差異;下表列出了第2天與第7天之間比較的P值(ANOVA,Tukey檢驗,P≤0.05,n=6±標準誤)。
由于關于低氧誘導HsAQP1表達的報道表明水通道蛋白介導的轉運過程在低氧條件下可能特別重要,我們檢測了煙草的轉錄水平。低氧導致七天后低氧和通氣良好植物的葉片和所有根段中ATP水平相似。結果表明,在經歷初始低氧脅迫后,植物可能獲得了足夠的氧氣來支持有氧呼吸,因為低氧植物外觀健康翠綠,并且沒有表現出缺氧癥或其他可見的氧氣缺乏跡象。雖然某些植物對根低氧的抵抗可以通過形成特殊的通氣結構(如通氣組織)來增加向根細胞的氧氣供應來解釋,但在沒有明顯結構變化的情況下植物對低氧抵抗的過程仍然不清楚。在我們的研究中,暴露于根低氧的植物根和莖中沒有出現可指示改善氧氣輸送的結構特征。因此,NtPIP1;3轉錄水平的增加可能是促進根通氣的重要因素之一,類似于低氧人體組織中HsAQP1轉錄水平的增加。顯然,孔介導的氧氣轉運與低氧之間的聯系值得進一步關注。
總之,我們的結果表明,所研究的一些植物和人類水通道蛋白可能參與氧氣轉運。異源表達這些水通道蛋白的酵母細胞在液體培養中保持了更高的氧氣吸收速率,并且煙草植物在其根遭受低氧條件后,推定的氧氣轉運水通道蛋白急劇增加。這些氧氣轉運水通道蛋白在七天低氧處理后的增加伴隨著低氧根尖段ATP水平的增加。研究結果支持了孔介導的氧氣轉運的功能意義應受到更多關注的觀點。
 相關新聞
相關新聞