熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
3.結果與討論
3.1.初始條件
維戈河口是一個非常多變和動態的海灣,具有明顯的季節性模式,包括兩個不同的時期:涌升季節(3月至9月),以低溫和高鹽度表層水為特征,葉綠素a濃度高,以微型浮游生物(>20微米)為主;下沉流季節(10月至3月),水溫較高,葉綠素a濃度低,小型浮游生物占浮游植物現存量的主導地位。涌升條件在9月的時間序列(時間序列1和時間序列2)期間盛行,其特征是時間序列1期間的強北風和時間序列2期間的風力減弱。相對較高的表層溫度和鹽度表明處于涌升減弱期。采樣日的初始葉綠素a濃度很高(時間序列1和時間序列2分別高于6.7和10微克葉綠素a/升),是該時期的特征。經過樣品三天的預處理后,時間序列2初始時間的葉綠素a降至0.45微克葉綠素a/升。盡管在時間序列1和時間序列2期間未進行浮游植物計數,但在高葉綠素a、有利的涌升期,微型浮游生物(>20微米)通常主導浮游植物群落,這與時間序列2預處理后濃度的顯著下降一致。
相反,秋冬條件在時間序列3-時間序列5期間盛行,其特征是12月和1月采樣期間的南風/西風。由于采樣期間經歷強降雨和相關的高大陸徑流,發現了相對較低的溫度值(范圍11-12°C)和低鹽度值。葉綠素a濃度達到下沉流季節的典型值,范圍在0.3-0.55微克/升。最豐富的浮游植物類群是鞭毛藻和甲藻(分別占總豐度的42-78%和21-47%),盡管硅藻和纖毛蟲也存在,但比例較小(分別為0.1-12%和0.1-7%)。時間序列3和時間序列5初始樣品中較低的葉綠素a濃度和硅藻的稀少存在源于3天的沉降過程。浮游植物的初始豐度從12月(時間序列3)的約2 x 10^9細胞/升下降到2月初(時間序列5)的約0.2 x 10^9細胞/升。相反,12月發現最低的初始細菌豐度(0.3x10^6細胞/升),在1月至2月達到1.6x10^6細胞/升,這是每年這個時間的特征。
這種自然和人為誘導的初始條件變異性使我們能夠在更廣泛的浮游生物群落范圍內測試偏差。
3.2.時間序列實驗
3.2.1.葉綠素a和微型浮游生物豐度
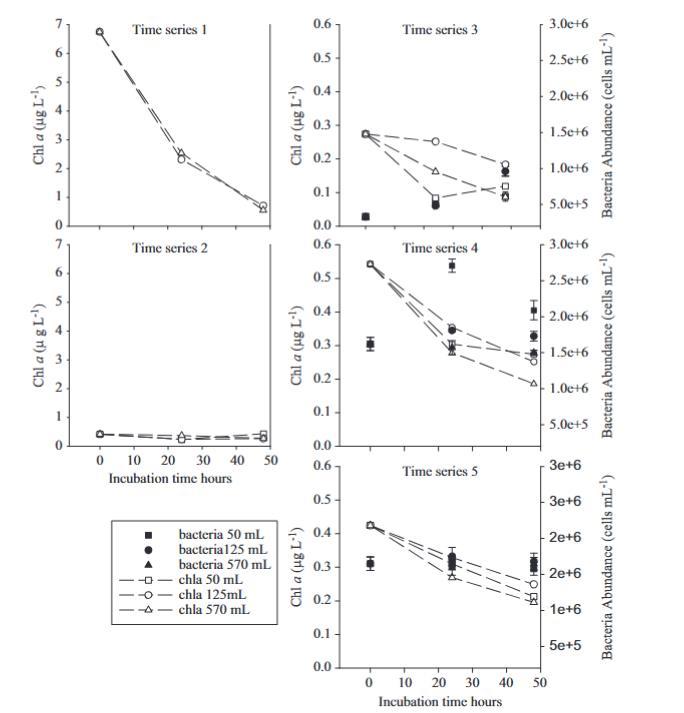
圖2.不同時間序列實驗中的葉綠素a濃度和細菌豐度。深色符號(■●▲)對應細菌豐度,而白色符號(□△)對應葉綠素a濃度。誤差棒代表1個標準差。
在所有培養過程中,除了時間序列3,葉綠素a濃度在三種瓶子體積中以相似的方式下降(圖2)。時間序列1的初始葉綠素a濃度非常高,對應于后涌升情況,經歷了急劇下降(62-65%),這在其他實驗中未見(約32%)。與秋季實驗一樣,在冬季,未經處理的實驗(時間序列4)中葉綠素a的下降幅度高于經過處理的實驗(時間序列3,時間序列5)。
冬季實驗中微型浮游植物細胞數量反映了與葉綠素a在三種瓶子體積中相似的趨勢(圖3),除了時間序列5,其中細胞數量在48小時后增加了兩倍。微型浮游植物總是以鞭毛藻和甲藻為主,但在三個冬季時間序列之間觀察到群落結構存在差異。硅藻僅存在于未經處理的時間序列4實驗中(零時占總豐度的12%),并且硅藻和甲藻的豐度在實驗末期均減少。在時間序列3和時間序列5中,甲藻對浮游植物群落的貢獻在實驗末期增加;而鞭毛藻的豐度在時間序列3中下降,在時間序列5中增加。
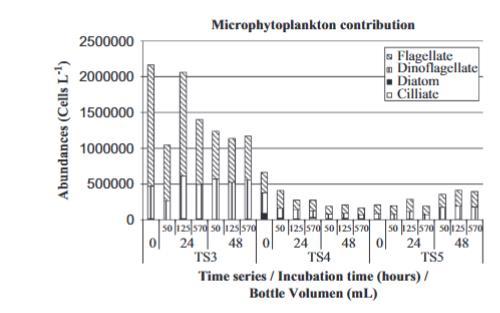
圖3.不同時間序列實驗中最豐富的微型浮游植物類群的貢獻。
3.2.2.細菌豐度
在冬季實驗中,初始時間的細菌豐度范圍從0.3到1.6 x 10^9細胞/升,12月最低,1月至2月(時間序列4和時間序列5)最高。在每個時間序列中,三種瓶子大小的細菌遵循幾乎相同的趨勢,除了時間序列4,其中50毫升瓶子在培養24小時后顯示出更高的豐度(圖2)。
只有在時間序列3中,細菌從非常低的初始數量增加到培養開始時的兩倍。這是唯一一個鞭毛藻初始豐度非常高并在時間序列期間下降的實驗,這可能表明鞭毛藻的噬菌作用在控制細菌豐度方面起作用。
總的來說,這五個實驗提供了一系列不同的浮游生物初始條件和演化過程。
3.2.3.群落呼吸速率
文獻調查表明,125毫升是暗群落呼吸培養最常用的瓶子體積,而瓶子培養的標準時間是24小時,因為驅動氧濃度變化的生理過程(生產和呼吸)對晝夜光周期敏感,因此只有測量24小時總初級生產/群落呼吸平衡才能準確代表群落的凈有機物生產。因此,我們在此呈現的結果分析將特別關注125毫升瓶子和24小時培養。
3.2.4.離散培養
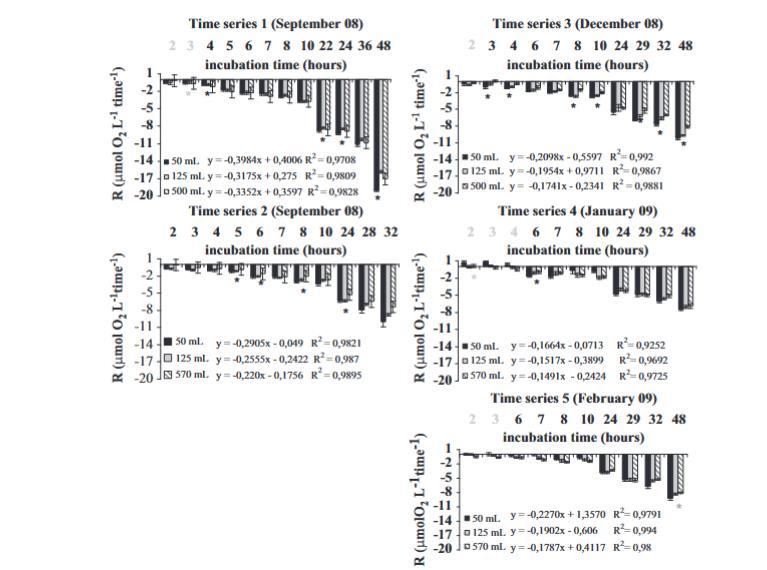
圖4.每種瓶子體積在不同培養期后的累積呼吸量。加粗的小時數表示介于最小和最大培養時間之間的時段。標記(*)表示某個瓶子存在統計學差異。回歸方程是從最小培養時間到最后一個培養時間計算的。請注意,x軸不是線性時間軸。
圖4顯示了三種大小瓶子在不同培養時間后的耗氧量。附錄1包含了每個培養期的累積呼吸速率和標準偏差。在檢驗任何假設之前,我們需要評估產生可靠呼吸數據的最小培養時間,從而確定每種時間序列技術的靈敏度。對于標準的125毫升瓶子體積,最小時間因實驗而異,范圍從9月的2小時到1月至2月的6小時(圖4和表1)。除時間序列1和時間序列5中的50毫升瓶子外,同一實驗中不同大小瓶子的最小時間差異相對較小。最小時間消耗的氧氣范圍在0.6到1.6微摩爾O2/升之間(時間序列1-時間序列2和時間序列5)。觀察到24小時呼吸速率與最小時間之間存在顯著的負相關關系。
對于長于最小時間的培養,僅在131次培養中的30次中發現三種不同瓶子體積之間存在顯著差異(圖4)。差異通常表現為570毫升瓶子的暗群落呼吸速率較低(47%),與其他兩種瓶子體積相比(與50毫升瓶子有23次顯著差異,與125毫升瓶子僅有7次顯著差異)。125毫升瓶子與其他兩種瓶子大小之間暗群落呼吸速率的統計學顯著差異平均幅度小于14%。
在標準的24小時培養中,僅在時間序列1和時間序列2中發現顯著差異(表1),平均約占暗群落呼吸速率的12.5%。如果我們比較三種瓶子體積的耗氧量對時間回歸線的斜率(從最小時間到最大時間),顯著差異也僅出現在時間序列1和時間序列2中。
| 實驗 | 實驗體積(mL) | 溫克勒方法 | 呼吸計R 24小時 μmolO? L?1 天?1 | p值斜率差異 (溫克勒125 vs 呼吸計) | |||||
| R 24小時 μmolO? L?1 天?1 | 標準誤 | 最小時間(小時) | 最大時間(小時) | R 24小時差異 | 斜率差異 | ||||
| 時間序列1 | 50 | 9.27 | 0.17 | 2 | >570>125 | ≠125≠570 | |||
| 125 | 8.46 | 0.15 | 4 | ≠50 | |||||
| 570 | 8.86 | 0.17 | 4 | 36 | >125 | ≠50 | |||
| 時間序列2* | 50 | 6.29 | 0.25 | 2 | >570 | ≠125≠570 | |||
| 125 | 6.29 | 0.18 | ≠50≠570 | ||||||
| 570 | 5.18 | 0.18 | 3 | ≠125≠50 | |||||
| 時間序列3* | 50 | 5.55 | 0.37 | 5.04 | 0.41 | ||||
| 125 | 5.28 | 0.61 | |||||||
| 570 | 4.84 | 0.16 | 4 | ||||||
| 時間序列4 | 50 | 4.78 | 0.27 | 6 | 29 | 4.71 | 0.71 | ||
| 125 | 3.99 | 0.28 | 6 | ||||||
| 570 | 4.30 | 0.28 | |||||||
| 時間序列5* | 50 | 3.82 | 0.23 | 32 | 4.22 | 0.14 | |||
| 125 | 3.78 | 0.26 | 10 | 29 | |||||
| 570 | 3.32 | 0.2 | |||||||
這些結果表明我們可以拒絕前兩個假設:"瓶子效應"不會系統性地導致呼吸估算準確度誤差,并且125毫升培養對于維戈河口的暗群落呼吸測量是足夠的。
 相關新聞
相關新聞